
Lo que nuestros genes (y los de nuestros antepasados) cuentan sobre nosotros
Imagina que guardas una carta escrita hace diez mil años. No en papel, que no existía, sino en ti mismo. En cada célula de tu cuerpo hay un texto antiquísimo que narra de dónde vino la gente que acabó, generación tras generación, convirtiéndose en ti. Hasta hace poco no teníamos manera de leerlo, pero ahora sí.
Llevo años curioseando y leyendo artículos, algún libro y cosas varias sobre expertos que analizan el ADN de restos humanos de miles de años de antigüedad (arqueogenómica, creo que se llama) y lo que he encontrado me parece fascinante. No soy científico: soy alguien que lee estas cosas con curiosidad y voy a intentar contar aquí un poco sin distorsionarlas demasiado. Todo lo que he escrito aquí tiene detrás estudios publicados en revistas especializadas, pero la interpretación es mía, y asumo que puede tener imprecisiones. Sólo espero que os interese tanto como a mí.
Homo Sapiens: una especie joven, de origen y entremezclada
Lo primero que hay que saber es que, según dicen los genetistas, los seres humanos somos, una especie extraordinariamente homogénea en términos evolutivos. Esto quiere decir que, comparados con otras especies, hay muy pocas diferencias en el ADN entre dos individuos. Toda la variabilidad genética humana actual es menor que la que existe entre dos grupos de chimpancés en la misma selva africana, que tienen el doble o hasta el triple de variabilidad genética que el Homo Sapiens. Por tanto, nuestras diferencias genéticas son proporcionalmente muy pequeñas comparadas con las de nuestros parientes más cercanos. Somos una especie joven que lleva relativamente poco tiempo diferenciándose.
¿Por qué somos tan poco diversos? Hay quien cree que hubo un cuello de botella en algún momento entre 50.000-100.000 años (quizá la erupción del supervolcán Toba, en Sumatra, hace unos 74.000 años) donde la población de Homo Sapiens se redujo a pocos reduciendo claramente la variabilidad genética.
Y sin embargo, esa historia corta está llena de migraciones, encuentros y mezclas. Lo que la genética está revelando es que no hay ninguna población humana que no sea producto de múltiples oleadas de mestizaje. Ninguna. Incluso las más «aisladas» son resultado de al menos 3 grandes oleadas de poblaciones distintas.
La hipótesis de que el Homo Sapiens se originó en África está ampliamente aceptada y sostenida por la genética (todos los no africanos actuales compartimos un ancestro común y la diversidad genética decrece desde África subsahariana hacia el exterior) y por la paleontología (casi no se ha encontrado fósiles anteriores a 120.000 años fuera de África y se disparan a partir de hace 70.000 años). Y precisamente, la expansión fuera de África (llamada modelo «Out of Africa«) favoreció que cada grupo que se separaba y se asentaba en un nuevo territorio llevaba consigo una parte de la diversidad genética (esto se llama «efecto fundador«) y es el motivo por el que las poblaciones más alejadas de África son las menos diversas genéticamente.
Ojo, no estamos hablando de rasgos ni de razas. Los rasgos que usamos para clasificar a las personas visualmente (como el color de piel, el pelo, la forma de los ojos, la altura), al parecer, se deben a muy pocos genes y tienen que ver con la adaptación a condiciones naturales (el sol, el frío, la altitud…). El resto del genoma (el 99,9%) no tiene que ver con el aspecto físico y es tremendamente uniforme entre todos los humanos. Vamos, que según los genetistas, no existen «razas» humanas en el sentido genético. Llevamos unos 300.000 años como especie y sólo 65.000-70.000 fuera de África. No ha habido tiempo suficiente para divergir genéticamente de forma profunda.
Del modelo lineal que estudiamos en el colegio a un modelo entrelazado cada vez más complejo
El modelo lineal que estudiamos hace años:
Todos estudiamos en el colegio que la evolución desde el Australopitecos hasta el Homo Sapiens era una secuencia lineal:
Homo Habilis → Homo Erectus → Homo Neanderthalensis / Homo Sapiens
Según aquello, cada especie sustituía a la anterior, y que el Homo sapiens era el «producto final» de una evolución progresiva y bastante ordenada. Incluso, en algún momento al Homo Neanderthalensis se le denominó «Homo sapiens neanderthalensis» y a nosotros «Homo sapiens sapiens«, como si uno fuera evolución del otro. Después ya los neandertales se empezaron a considerar una rama lateral que simplemente se extinguió cuando llegamos nosotros, sin mezclarse. Y hoy sabemos que tampoco es así: que cada uno es una línea distinta (y que el Neardental es una variable concentrada en Europa y Oriente Medio), aunque también recientemente se ha descubierto que hubo una mínima hibridación.
El modelo entremezclado que conocemos ahora:
Gracias al trabajo de los genetistas (especialmente Pääbo que secuencio el genoma Neardental en 2010 y ganó el Nobel en 2022, ojito), hoy sabemos que Los europeos y asiáticos comparten entre un 1,8% y un 2,5% de ADN con los neandertales. Los africanos subsaharianos, no. Eso sólo podía significar una cosa: esa pequeña mezcla ocurrió después de la salida de África.
- Sapiens + neandertales → todos los no-africanos tenemos ~2% ADN neandertal.
- Sapiens + denisovanos → los melanesios y aborígenes australianos tienen hasta un 4-6% denisovano.
- Neandertales + denisovanos → también se mezclaron entre sí.
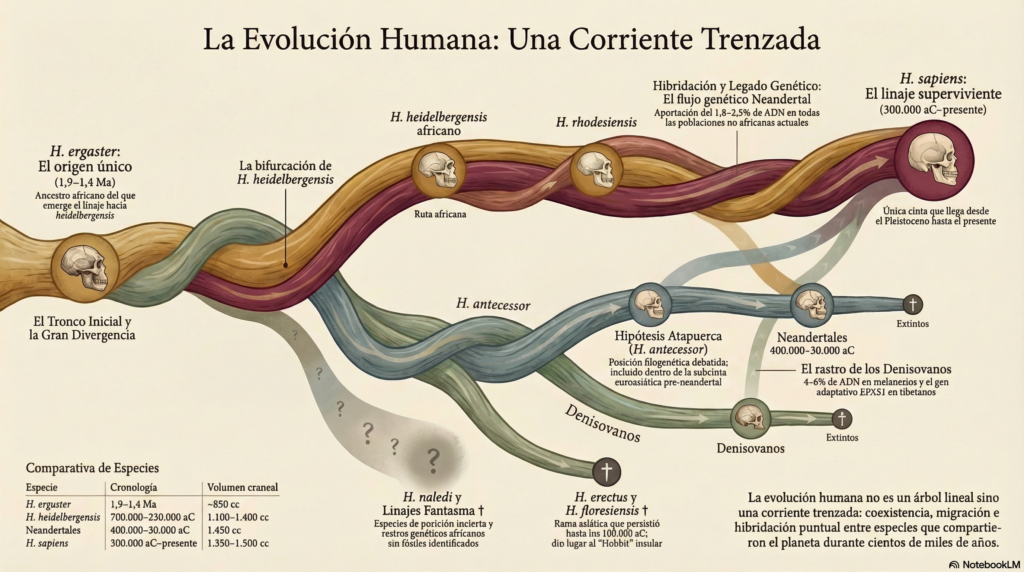
Pero no sólo convivieron y se mezclaron (muy poco) los Sapiens y Neardentales. Antes, muchos individuos del género Homo coexistieron en los mismos territorios, los cruces no fueron eventos puntuales, hay especies que se acaban considerando variaciones geográficas de la misma especie o, incluso, el ADN revela especies que se mezclaron con Sapiens o Neandertales pero de las que no tenemos ningún fósil. Cuanto más conocemos, más lioso es el diagrama y cada vez se parece más a un arbusto con ramas entrelazadas y menos a un árbol lineal, como se creía hasta hace poco.
Según el consenso científico actual, no descendemos ni del Homo Habilis ni del Homo Erectus. Sino que los descendientes del Australopithecus fueron 3 especies Homo que convivieron en África entre 2,4 y 1,6 millones de años aC:
- el Homo habilis (600-750 cc de cráneo).
- el Homo rudolfensis (750 cc de cráneo).
- el Homo ergaster (850 cc de cráneo), vivió de 1,9 a 1,4 millones de años aC.
El Homo sapiens desciende del Homo Ergaster que, a su vez, evolucionó:
- en Asia como Homo Erectus (900-1.200 cc de cráneo), que se extinguió en torno al 100.000 aC.
- en África como Homo Heidelbergensis (1.100-1.400 cc de cráneo) 700.000 a 230.000 aC.
El Homo sapiens desciende del Homo Heidelbergensis (el que se quedó en África) que, a su vez, evolucionó:
- en Eurasia como Homo Heidelbergensis europeo (aunque según la Escuela de Atapuerca, ya había llegado el Homo Ergaster como Homo Antecessor) que acabó evolucionando como:
- Homo Neanderthalensis en Europa y Oriente Próximo (1.450 cc de cráneo) de 400.000 a 30.000 aC.
- Homo Denisova en Siberia y Asia (con 1.400 cc de cráneo), debió extinguir se en torno al 50.000 aC.
- en África como Homo Rhodesiensis, Homo Heidelbergensis africano o también llamado Homo Sapiens arcaico (300.000 aC), que acabó evolucionando como:
- Homo Sapiens, nosotros (1.350 cc de cráneo) desde 200.000 aC hasta hoy.
Por tanto, la línea evolutiva correcta es:
Homo Ergaster → Homo Heidelbergensis → Homo Rhodesiensis → Homo Sapiens
En definitiva, los Homo Sapiens venimos de las ramas del género Homo que se fueron quedando en África, donde evolucionaron. Y ya después, salió de África (hipótesis ‘Out of Africa‘) en 2 oleadas: a) ~120.000 aC hacia Levante y no dejó descendencia detectable en poblaciones actuales; b) ~65.000–70.000 aC: la gran salida desde el cuerno de África hacia Arabia de la que procedemos todos los no africanos actuales.
Por curiosidad, el Homo Sapiens se encontró con los Neardentales en dos ocasiones: al salir de África en el 60.000 aC, en Oriente Próximo; y cuando el Sapiens entró en Europa en el 45.000 aC, comenzando el retroceso definitivo de los Neardentales hasta desaparecer en la Península Ibérica alrededor del 30.000 aC.
El ADN antiguo ha permitido cosas que complementan a los fósiles, como confirmar y estimar la fecha de la hibridación con los Neardentales o los Denisovanos (en este último caso sin tener casi ningún fósil). Y el gran nombre ha sido el ya citado Svante Pääbo (Nobel 2022).
Cómo leer genéticamente el pasado: Y-DNA y mtDNA
Antes de entrar en las grandes migraciones, creo que vale la pena explicar cómo funciona el sistema de rastreo que usa la genómica para reconstruir todo esto. Porque cuando empiezas a leer sobre el tema, aparecen constantemente dos conceptos: el cromosoma Y y el ADN mitocondrial, y entenderlos es clave. A mí es lo que más me cautivó para seguir indagando.
El cromosoma Y: la línea paterna (Y-DNA)
El cromosoma Y (Y-DNA) se hereda exclusivamente de padre a hijo. Un hombre tiene exactamente el mismo cromosoma Y que su padre, y el mismo que su abuelo paterno, su bisabuelo paterno… con pequeñas mutaciones que se van acumulando a lo largo del tiempo. Eso lo convierte en una herramienta ideal para rastrear la línea masculina a través de los siglos e identificar migraciones geográficas.
Cuando una mutación aparece en un hombre y se propaga por sus descendientes, se crea lo que se llama un haplogrupo: una familia de cromosomas Y que comparten esa mutación como marca de origen común. Los haplogrupos tienen nombres técnicos (R1b, I2, E-M81…) que al principio parecen jeroglíficos, pero que básicamente son apellidos genéticos de linajes masculinos.
CURIOSIDAD: El haplogrupo R1b es el más común entre los varones de Europa occidental y llegó con los pastores de la estepa hace unos 4.000-4.500 años. Antes de su llegada era prácticamente inexistente en Europa. Según lo que he leído, en la Península Ibérica se produjo una sustitución de casi el 100% de los cromosomas Y locales en apenas unos pocos siglos.
El ADN mitocondrial: la línea materna (mtDNA)
El ADN mitocondrial (mtDNA) funciona de forma análoga pero en sentido contrario: se hereda exclusivamente por vía materna. Una mujer transmite su mitocondrial a todos sus hijos, pero sólo las hijas lo pasan a la siguiente generación. Igual que el Y-DNA, acumula mutaciones que permiten identificar distintos linajes maternos y rastrear las migraciones geográficas.
Asimetría entre linaje materno y linaje paterno.
Una de las cosas más curiosas es que, como os podréis imaginar, no tienen por qué coincidir el ritmo de las migraciones del linaje femenino y del masculino. De hecho, los linajes maternos (mtDNA) son, en general, considerablemente más antiguos y conservadores que los paternos. ¿Por qué? pues porque los hombres se desplazaron más y las migraciones prehistóricas tuvieron con frecuencia un componente de sustitución masculina mucho mayor que femenino.
El caso vasco es principal ejemplo de la asimetría: resulta que el 80% de los varones vascos porta hoy el haplogrupo R1b (Y-DNA), asociado a la expansión indoeuropea en Europa occidental hace unos 4.500 años. Sin embargo, sus linajes maternos son los haplogrupos H1, U5b (mtDNA), lo que parece mostrar una continuidad en el linaje femenino con las poblaciones pre-indoeuropeas que habitaban la región. La conclusión que sacan los investigadores es que el legado cultural anterior a los indoeuropeos (como la lengua primitiva) sobrevivió, pero la línea masculina fue reemplazada casi por completo por la oleada indoeuropea (R1b alto).
UNA PREGUNTA: Si los vascos arcaicos tienen la misma proporción de R1b indoeuropeo que sus vecinos, ¿cómo conservaron una lengua pre-indoeuropea cuando nadie más en Europa occidental lo hizo? La respuesta más aceptada tiene que ver con el aislamiento geográfico y una endogamia prolongada que ralentizó los intercambios culturales posteriores. Por lo que hemos visto en el párrafo anterior, quizá fuera clave el linaje materno. Pero sigue siendo, en mi opinión, uno de los misterios más interesantes de la prehistoria europea.
Hoy R1b alcanza frecuencias altísimas en el extremo occidental de Europa:
| Región | Frecuencia R1b (Indoeuropeo) |
|---|---|
| Irlanda | ~80-85% |
| País Vasco | ~75-80% |
| Gales | ~75% |
| España en general | ~60-70% |
| Francia | ~60% |
| Italia norte | ~40% |
| Europa del este | ~20-30% |
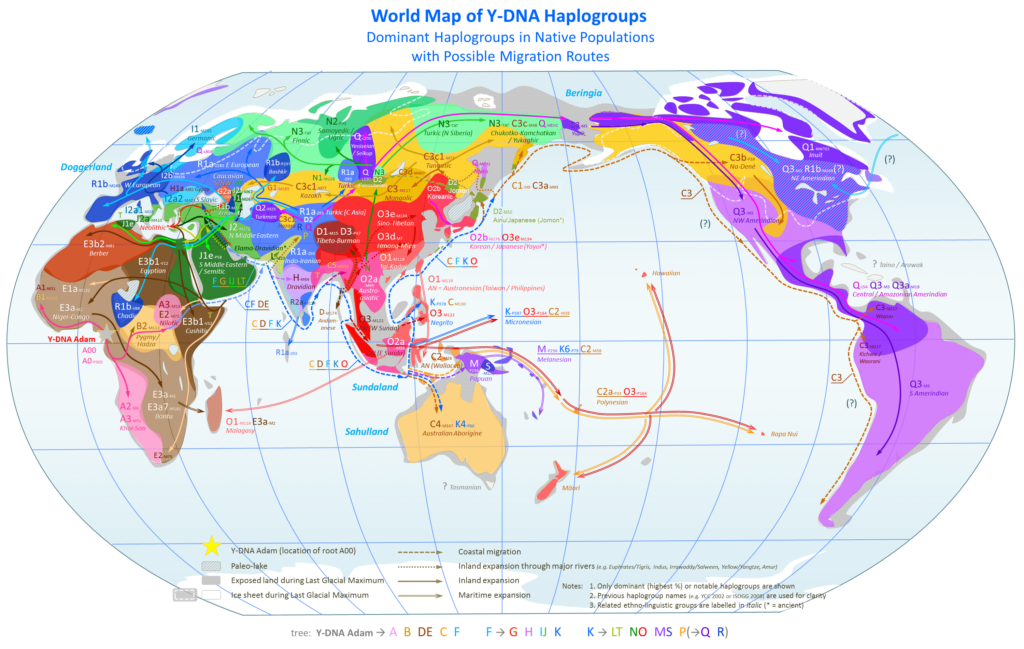
Hablaremos mucho de los haplogrupos (tanto mitocondriales como cromosómicos) porque, como en el mapa anterior, permiten visualizar las distintas migraciones del Homo Sapiens a partir del estudio genético. Cómo se movió el linaje materno (mtDNA) y cómo el linaje paterno (Y-DNA). Evidentemente, el mapa no es definitivo, sino que va cambiando a lo largo de los años, a medida que se descubren nuevas

El origen: Africa
Mientras la 3ª glaciación castigaba Europa con frío extremo y un clima árido y hostil, África mantuvo riqueza y variedad ecológica que permitió al ya citado el Homo heidelbergensis africano (o H. rhodesiensis) adaptarse y acumular genéticamente mutaciones durante cientos de miles de años gracias a los ciclos ecológicos africanos. Básicamente, el entorno no era estable sino que cambiaba constantemente con períodos húmedos, períodos secos, sabana, bosque, de nuevo sequía,…), lo que obligaba a adaptarse una y otra vez. En ese contexto, los individuos que sabían cooperar, comunicarse mejor y resolver problemas nuevos sobrevivían y tenían más descendencia que los que no. La inteligencia social, el lenguaje y la capacidad simbólica aparecieron poco a poco y resultaron útiles para sobrevivir. Así, hace unos 300.000 años, emergió el Homo Sapiens, cuyos restos más antiguos se han encontrado un yacimiento de Marruecos. No tengo mucha idea de las diferencias entre aquel y el H. Sapiens anatómicamente moderno, que aparece en África subsahariana en torno a 200.000 aC.
Desde el punto de vista del linaje materno (mtDNA):
El linaje mitocondrial más antiguo conocido es el haplogrupo L0, datado en torno a 150.000 aC en el sur de África. Los haplogrupos mitocondriales L1 y L2 se quedaron en África y son los predominantes en África subsahariana occidental y central hoy. Sin embargo, L3 surgió en el este de África (actual Sudán y Etiopía) entre 80.000 y 70.000 aC.
Desde el punto de vista del linaje parterno (Y-DNA):
El primer linaje cromosómico (Y-DNA) es el haplogrupo A (en el Noroeste de África y descendiente del “Adán cromosómico”). De él surgen: A2 en el sur de África, A3 en el este BT que, a su vez, se subdivide en B (sólo en África) y CT surgido entre 100.000- 70.000 aC en el Este de África (Etiopía-Sudán)
Durante la cuarta y última glaciación (el Würm o Edad de Hielo, entre 115.000 y 11.700 aC) el nivel del mar descendió considerablemente, abriendo pasos hoy sumergidos. Gracias al análisis genético, los científicos consideran que, en algún momento hace 65.000-67.000 años, un grupo de Homo Sapiens portadores del haplogrupo mitocondrial L3 y del haplogrupo cromosómico CT cruzó el estrecho de Bab-el-Mandeb, que separa el Cuerno de África de la península arábiga, y siguió la costa sur hasta alcanzar el Valle del Indo. Fue la segunda oleada a través de la llamada ruta sur, la gran migración fundacional de toda la humanidad fuera de África, ya que todos los no africanos descendemos del L3 (mtDNA) y del CT (Y-DNA).
Fuera de África, el boom demográfico desde Indostán
El pequeño grupo que salió de África encontró en el actual Oriente Medio y el valle del Indo territorios fértiles, vacíos de competidores humanos y con recursos abundantes, que actuó como acelerador demográfico: la pequeña población fundadora se expandió rápidamente y comenzó a ramificarse: a) el linaje materno L3 se dividió en sus dos grandes ramas: M, que siguió la ruta costera hacia el sureste asiático y Oceanía, y N, que se quedó en el Indostán y comenzó a expandirse hacia el norte y el oeste; b) y simultáneamente en el linaje paterno CT hacía exactamente lo mismo: su rama D acompañó al M hacia oriente, mientras que CF —y de él F— se quedó y siguió ramificándose hacia el oeste.
La rama que acabaría llegando a Europa es en ambos casos la occidental. En el linaje materno, N derivó en R, que avanzó hacia Oriente Medio y el Cáucaso, donde dio lugar a HV y U, los haplogrupos maternos de los primeros europeos. En el linaje paterno, F generó sucesivamente I (los cazadores-recolectores que llegaron a Europa hace 46.000 aC), G (los agricultores neolíticos de Anatolia, 6.000 aC) y R1b (la oleada de indoeuropeos de las estepas al norte del Caspio, 3.000 aC). Dos linajes, materno y paterno, que narran el mismo viaje: de un estrecho africano a Europa, en oleadas sucesivas a lo largo de decenas de miles de años.
Llegada a Europa
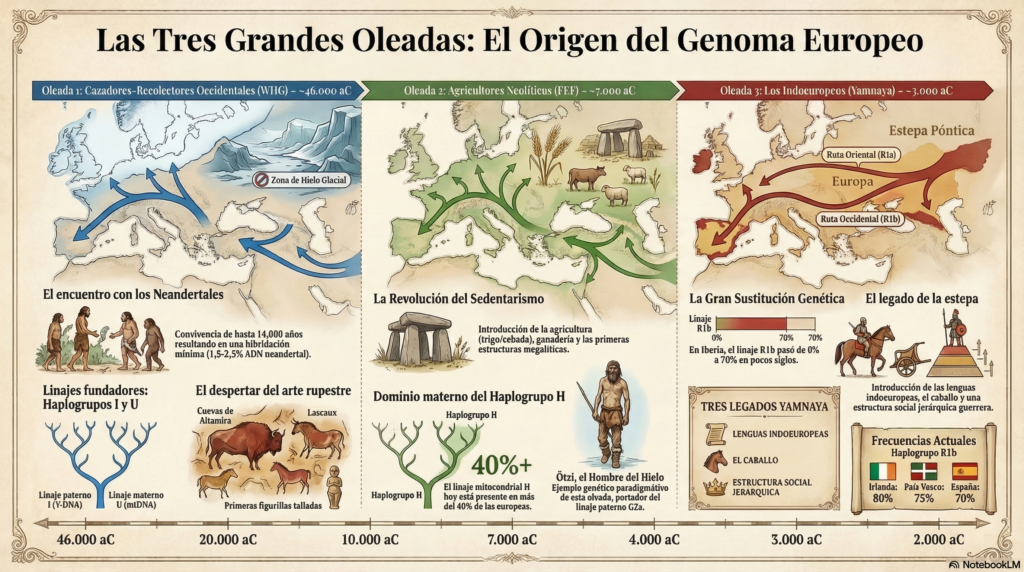
Por tanto, tres oleadas de llegada a Europa:
1ª Oleada: Los «cazadores-recolectores occidentales» (‘WHG’).
Los ‘WHG’ tenían linajes paterno I (Y-DNA) y materno U (mtDNA) que llegaron a Europa hace 46.000 aC desde Oriente Próximo y el Cáucaso: encontraron un continente prácticamente vacío pero helado en la Edad de Hielo con el norte impracticable y, por ello, se concentraron en las franjas del sur (la Península Ibérica, el sur de Francia, los Balcanes). Pintaron Altamira y Lascaux, fabricaron las primeras figurillas y enterraron a sus muertos con ocre y adornos. Y, como hemos dicho, allí se encontraron con los neandertales que llevaban más de 300.000 años en Europa. La convivencia duró entre 3.000 y 14.000 años según la región, con quienes convivieron varios miles de años y se mezclaron mínimamente (según los citados trabajos de Pääbo, todos los no-africanos actuales portamos entre un 1,8% y un 2,5% de ADN neandertal) antes de desplazarlos definitivamente.
2ª Oleada: Los «agricultores de la revolución neolitica» (‘EEF’).
Los agricultores neolíticos tenían el linaje maternolineal HV (mtDNA) y sus derivados, especialmente H (hoy el más frecuente en Europa, presente en más del 40% de las mujeres europeas actuales) y el linaje masculino G (Y-DNA), llegaron a Europa en el 7.000-6.000 aC y se expandieron desde Anatolia a través de 2 rutas: una costera mediterránea y otra por el interior de los Balcanes hacia el centro del continente. Trajeron la agricultura (trigo, cebada…) la ganaderia (ovejas, cabras…), la cerámica, los poblados estables y primeras estructuras megalíticas. Su expansión demográfica fue muy superior. En las zonas de mayor fertilidad agrícola los agricultores se impusieron con rapidez, desplazando a los cazadores-recolectores originales.
3ª Oleada: Los indoeuropeos.
Los hoy llamados ‘indoeuropeos’ eran un grupo de pastores seminómadas que había desarrollado una combinación de ventajas como el caballo domesticado, el carro y una organización guerrera y jerárquica. Genéticamente, eran un pueblo mestizo surgido de la mezcla entre los cazadores-recolectores de las estepas orientales («EHG», Eastern Hunter-Gatherers) y los cazadores-recolectores del Cáucaso («CHG») y portaron el linaje paterno R1b (Y-DNA) y los maternos H, U5 y T (mtDNA) que ya existían. Llegaron a Europa en el 3.000-2.500 aC desde las estepa Póntica (al norte del mar Negro y el Caspio, el actual sur de Ucrania, el norte del Cáucaso y el suroeste de Kazajistán) con dos rutas: los portadores de R1a llegaron hasta la India, Persia y Asia central, llevando consigo las lenguas que darían lugar al sánscrito, el persa y las lenguas eslavas y bálticas y los portadores del R1b hacia el oeste, llegando a la Península Ibérica alrededor del 2.500-2.200 aC, asociada a la cultura del vaso campaniforme.
El haplogrupo paterno G2a de los agricultores neolíticos prácticamente desapareció en una o dos generaciones, sustituido de forma masiva por R1b. La velocidad y la magnitud de la sustitución no tienen parangón en el registro genético europeo. En la Península Ibérica, por ejemplo, el R1b pasa de ser inexistente antes de 2.500 aC a representar más del 70% de los linajes masculinos apenas unos siglos después. Eso sugiere un proceso de imposición violenta o jerárquica: los varones yamnaya se reprodujeron masivamente con mujeres locales. Llegaron a Europa con tres legados que perduran hasta hoy: 1) Las lenguas indoeuropeas: el español, el inglés, el francés, el alemán, el griego, el ruso, el hindi; 2) El caballo y la movilidad; y 3) Una nueva estructura social. El R1b es hoy el haplogrupo paterno dominante en Europa occidental: 80% en Irlanda, 75% en el País Vasco, 70% en España, 60% en Francia, como ya hemos visto. La mayoría de europeos occidentales que se se hacen una prueba de ADN acaban encontrando R1b en su linaje paterno.
Lecturas:
— «La especie elegida» Arsuaga, J.L. & Martínez, I. (1998). Me encantó. Quizá pensado para el gran público sobre la evolución humana. Explica exactamente la secuencia H. Ergaster → H. antecessor → H. heidelbergensis europeo → Neandertales, desde la perspectiva de la Escuela de Atapuerca.
— «Atapuerca y la evolución humana«. Arsuaga, J.L. & Martínez, I. (2004). Fundació La Caixa. Para el contexto de los homínidos europeos antes del sapiens.